A Rubric of the Revised Gnostic Mass
For the Private Working Couple
Issued by Order of the Acting Council of Elders
Gnostic Church of L.V.X. Member A.M.H.R.
– Of the Breviary of the G.C.L. —
Do what thou wilt shall be the whole of the Law.
Let the temple be arranged with the bed in the east and in the west, a ewer of water and salt. The Priest begins in the north as the censor is placed in the south. In the center is the altar attended by the Priestess and displaying an image of Horus, Liber Al vel Legis, dagger, bell and oil.
The Priestess is dressed in a blue robe with silver vestments and a field of stars about her head. The Priest is dressed in a white robe with red vestments and the serpent crown.
The Room is banished and sealed by the Priest.
PRIESTESS:
Silence! The moon ceaseth (her motion),
That also was sweet
In the air, in the air, in the air!
Who Will shall attain! Who Will shall attain
By the Moon, and by Myself, and by the Angel of the Lord!Now Silence ceaseth
And the moon waxeth sweet;
(It is the hour of) Initiation, Initiation, Initiation.
The kiss of Isis is honeyed;
My own Will is ended,
For Will hath attained.
PRIEST:
Behold the lion child swimmeth (in the heaven)
And the moon reeleth:
(It is) Thou! (It is) Thou! (It is) Thou!
Triumph; the Will that staggered
Before Ra Hoor Khuit!—Hadit!—Nuit!To the God OAI
Be praise
In the end and the beginning!
And may none fall
Who Will attain
The Sword, the Balances, the Crown!
PRIESTESS:
I am the harlot that shaketh Death.
This shaking giveth the Peace of Satiate Lust.
Immortality jetteth from my skull,
And music from my vulva.
Immortality jetteth from my vulva also,
For my Whoredom is a sweet scent like a seven stringed instrument,
Played unto God the Invisible, the all ruler,
That goeth along giving the shrill scream of orgasm.
PRIEST:
I am a man among men.
The Priestess takes an ewer of Water in one hand and Salt in the other. She then sprinkles Salt into the Water and mixes it with her hand while proclaiming:
Let the salt of Earth admonish the Water to bear the virtue of the Great Sea.
Mother, be thou adored!
The Priestess then rises and turns; taking some Water and makes three crosses over the Priests forehead, breast and body while declaring:
Be the Priest, pure of body and soul!
The Priestess then takes the open censor in one hand and incense in the other. She places incense on the hot brick while proclaiming:
Let the Fire and the Air make sweet the world!
Father, be thou adored!
The Priestess then takes the censor, making three crosses as before over the Priest while declaring:
Be the Priest, fervent of body and soul!
The Priestess then kneels, taking the Lance between her open hands, running them up and down upon the shaft very gently until a full and firm erection is garnered by the Priest and culminating at the end of the last stroke with the flinging of her hands to the heavens and proclaiming:
Be the Lord present amongst us!
The Priest then invokes:
Thee therefore whom we adore we also invoke. By the power of the lifted Lance!
The Priest takes the Priestess by her right hand with his left, keeping the Lance raised. He then proclaims:
I Priest and King greet thee and meet thee.
He raises her to her feet and states while escorting her to the foot of the bed:
I ascend with thee to the summit of the earth wherein you are enthroned and adored.
The PRIEST sprinkles the Priestess, making five crosses, forehead, shoulders, and thighs.
The PRIEST takes the censer and makes five crosses, as before.
Then, with direct eye contact between the couple:
The PRIEST:
O circle of Stars whereof our Father is but the younger brother, marvel beyond imagination, soul of infinite space, before whom Time is Ashamed, the mind bewildered, and the understanding dark, not unto Thee may we attain, unless Thine image be Love. Therefore by seed and root and stem and bud and leaf and flower and fruit do we invoke Thee.
Then the priest answered & said unto the Queen of Space, kissing her lovely brows, and the dew of her light bathing his whole body in a sweet-smelling perfume of sweat; O Nuit, continuous one of Heaven, let it be ever thus; that men speak not of thee as One but as None; and let them speak not of thee at all, since thou art continuous.
The PRIESTESS:
But to love me is better than all things; if under the night-stars in the desert thou presently burnest mine incense before me, invoking me with a pure heart, and the serpent flame therein, thou shalt come a little to lie in my bosom. For one kiss wilt thou then be willing to give all; but whoso gives one particle of dust shall lose all in that hour. Ye shall gather goods and store of women and spices; ye shall wear rich jewels; ye shall exceed the nations of the earth in splendour and pride; but always in the love of me, and so shall ye come to my joy. I charge you earnestly to come before me in a single robe, and covered with a rich head-dress. I love you! I yearn to you! Pale or purple, veiled or voluptuous, I who am all pleasure and purple, and drunkenness of the innermost sense, desire you. Put on the wings, and arouse the coiled splendour within you: come unto me! To me! To me! Sing the rapturous love-song unto me! Burn to me perfumes! Wear to me jewels! Drink to me, for I love you! I love you. I am the blue-lidded daughter of sunset; I am the naked brilliance of the voluptuous night-sky. To me! To me!
The PRIEST:
O secret of secrets that art hidden in the being of all that lives, not Thee do we adore, for that which adoreth is also Thou. Thou art That, and That am I.
I am the flame that burns in every heart of man, and in the core of every star. I am Life, and the giver of Life; yet therefore is the knowledge of me the knowledge of death. I am alone; there is no God where I am.
Thou that art One, our Lord in the Universe the Sun, our Lord in ourselves whose name is Mystery of Mystery, uttermost being whose radiance enlightening the worlds is also the breath that maketh every God even and Death to tremble before Thee. By the Sign of Light appear Thou glorious upon the throne of the Sun. Make open the path of creation and of intelligence between us and our minds. Enlighten our understanding. Encourage our hearts. Let thy light crystallize itself in our blood, fulfilling us of Resurrection.
Tuf ur biu
bi a’a chefu
Dudu nur af an nuteru.
The PRIESTESS:
There is no law beyond Do what thou wilt.
The PRIEST parts the veil with his lance.
Initiate sex; female superior is best as per the Riddle of the Sphinx. Upon penetration, the couple should alternately say the anthem between deep sensual kisses (stirring up the salivary glands) as follows:
Beast:
Pater et Mater unus deus Ararita.
Babalon:
Mater et Filius unus deus Ararita.
Beast:
Filius et Filia unus deus Ararita.
Babalon:
Filia et Pater unus deus Ararita.
They should fix their minds on the image of Baphomet as the Priest’s hands stimulate the various Priestess’ chakkras from the vital organs with endocrinal secretions (liver, kidneys, et al) and the spine itself, bringing the energy down from the Ajna into the Manipura where the female is focusing, while her hands are pressed over his Sahasrara. The male is focused on the ejaculant and the bud-will, bringing the heat from the hands of the Priestess, down from the Sahasrara and into the Mulhadhara.
When one partner is getting close to orgasm he or she should ‘Ararita’ as a que to their partner that they are nearing orgasm. If the other is not intoning this with them, they should pull back and resume the chant of the anthem. In this way, they may gauge their levels of excitement so that they may time their simultaneous orgasm with joint invocation of Ararita. The peak moment should then cause them to simultaneously shriek ‘Hriliu’ (meaning “I am he/she”) as the bud-will is poured into the ecstatic cup.
This may ultimately be consumed and shared orally by the Priest with the Priestess as they embrace in an energized egg of spirit; each visualizing the energy surrounding them and going through their united bodies in a tight embrace, their whole bodies being wrapped around each other.
Field Theory on Germination
The actual fertilization of the Egg is the working theory of this operation, and then to bring forth the egg from the vagina. But to get to this point, we need to examine what’s happening physically with the woman in order to produce the Egg. The essential scenario involves getting the Sperm to the Ovum and the Ovum must accept the Sperm. In the normal operation involving physical pregnancy, the womb is then depended upon to accept the fertilized Ovum for implantation in the Uterine Wall. For this, the Ovum is propelled downward by contractions of the fallopian tubes and by ciliary currents in the walls of the Womb. And its interesting to note that the fallopian tubes look like horns over the womb (symbolic of the ‘Belly of the Beast’), suggesting the image of the horned-god Pan, also known as Baphomet.
The consumption of the Egg fertilized with the bud-will captures the energy as a living force in the nature of this special Eucharist. But to get the Egg to the fallopian tube, ovulation needs to occur. This of course, happens during the first fortnight of the lunar cycle but can also be brought about through the sexual excitement of the woman. This is subsequently dependent upon how far the woman is involved in her arousal and her previous sexuality, including attitudes about her orgasm and on her womb contractions, how far down the womb reaches in orgasm and how active the cervix is. It is therefore, a good idea for the woman to map her cycle from month to month and to keep a dream diary as preparation for the operation.
Havelock Ellis notes that it may be possible that after menopause, an infinitesimal trace of ovarine may exist in order to preserve a woman’s femininity; just not enough to produce ovulation and menstruation. This is one avenue of research that also seems intriguing. It just doesn’t make sense that a woman should be cut out from the opportunity of this operation at as early an age as thirty-five. One avenue to look upon may be in the woman’s arousal and other attitudes as mentioned above. In this way, either the production of ovarine is increased by her arousal and excitement, enough so that even ovulation becomes possible or maybe even the ovarine is more easily captured in the Kteis.
1. Pre-ovulatory Phase
Ovarian follicles ripen & mature
The Egg grows in the Ovary in a little capsule called the follicle. The follicles are stimulated to grow by the ‘Follicle Stimulating Hormone’ (FSH) generated by the pituitary gland in the hypothalamus; associated with the autonomic nervous system. The hypothalamus is in effect, the unconscious mind as it is the leader of the endocrine system that orchestrates the development of the body, responding to the flowing blood in the body.
2. Ovulatory Phase (Culmination)
The ripe Egg is expelled from the ovary and the ‘Corpus Luteum’ ripens
The FSH stimulates the follicles by acting as a creative rash on the surface of the ovary; similar to putting sand in an oyster. The growth of the follicle produces oestrogen that spills into the blood going back to the pituitary gland and causing it to secretes the ‘Luteinizing (Yellowing) Hormone’, which causes one follicle to burst and release an egg, tube as it reaches for the Egg, leaving the fallopian tube to grow larger and turn yellow in color. The follicle is now referred to as the ‘Corpus Leutium’ or ‘Yellow Body’, which can now secrete progesterone and oestrogen; operating as a ductless gland.
3. Pre-menstrual Phase
Oestrogen & progesterone hormone levels drop
The progesterone stimulates the lining of the womb, causing it to thicken in preparation for a possible pregnancy. And when the progesterone reaches the pituatiry galnd, it stops the production of LH, causing the Corpus Leutium to die, should the ovum bot become fertilized. This leads to menstruation.
4. Menstrual Phase (Nadir)
Womb-lining shed
For but a particularly short moment linked with ovulation, the Gluten, normally acidic and destructive to the Sperm, becomes alkaline and conducive to the journey that the sperm must make in the womb. Yet at certain times, these juices thicken, making it more difficult for the Sperm to swim through the cervix. Do the ovaries produce more than one Egg and do they upon non-fertilization, fall through the cervix? If so, then the sperm can still fertilize the Egg without it being implanted onto the uterine wall. Or can the uterine wall be induced to expel it? Is this what was revealed to Crowley in the Amalantra working when he was told by the Wizard, “It’s all in the Egg”?
I see the egg on the pylon. I ask who the egg belongs to. The wizard shows me a hen laying an egg and says
“Whose egg is this?’’
Richard & Iona Miller in their essay, An Application of the Holographic Concept of Reality, state the following:
“Researchers have found that at the moment of ovulation there is a definite shift in the electrical fields of the body of the woman. The membrane in the follicle bursts and the egg passes down the fallopian tubes. As a side note, we feel that the phases of the moon quite probably influence the permeability of the membrane in the follicle, making it more likely that the egg will pass down the fallopian tubes at certain periods of time. The sperm is negative with respect to the egg. When the sperm and the egg unite, the membrane around the egg becomes hyper-polarized. It is at this moemt6n that the electromagnetic entity is formed.”
And for our purposes here, we can consider the speculative idea that Amalantra is one of the fabled Nephilim or Anunaki. These Nephilim were called ‘Sons of God’ and the Bible records them as having copulated with the ‘Daughters of Men’. The most ancient religions introduced the idea of a dual-natured or ‘bi-une’ god with male and female attributes. The idea of a single monotheist male god occurs much later in human history. Note the waters in Genesis are separated into the firmament above and the firmament below. The Starry Gnosis builds the sky into the ‘Egg’ that encompasses the whole sky with the Earth as its seed.
The Pharoah was said to have the power of rule only when he physically sat on the ‘Throne of Horus’. And the throne was also referred to being the Chariot of the Queen. Indeed, the heir to the throne of Egypt was produced by the child or the union of the son and the daughter. This not only tells us that the line of succession was matrilineal. But leads us to wonder why this particular bloodline was considered to be so important. For this, it gets particularly interesting in light of the scientific knowledge that we have of Mitochondrial DNA. Nuclear DNA is that formed in the zygote by the union of Nuclear DNA from both the sperm and the ova. However, Mitochondrial DNA is without the nucleus of the cell and is not penetrated by the sperm into the ova. Therefore, it is the mother’s Mitochondrial DNA that is transferred solely to the child. On rare occasions, the seminal Mitochondrial DNA has penetrated and altered the bloodline, but this is really the exception to the rule.
Regarding mithochondria, these "powerhouses" of the cell are bacteria which form a symbiotic relationship with eukaryotic (cells with a true nucleus) cells. That's why they have DNA. The reason inheritance is maternal is because mitochondria are found in the cytoplasm (and therefore outside the nucleus) and since the sperm carry almost no cytoplasm, the mitochondria in the offspring all come from the cytoplasm in the egg which is all maternal origin. Mitochondria reproduce independently in the cytoplasm and the cell does not have to divide to produce more mitochondria. There are numerous examples of eukaryotic cells (such as amoebae) that have what are known as endosymbiants in their cytoplasm. These are actually bacteria that have not exactly evolved into mitochondria yet. If you separate these endosymbiants from the host cell, they can live independently. However, since mitochondria are so specialized, a portion of the DNA of the mitochondria had been incorporated into the nucleus of the host cell and so it cannot live outside its host. There are still eukaryotic cells that do not have mitochondria. An example is the intestinal parasite Giardia Lamblia.
At the coronation of the Pharoah, he was anointed with that fat of the historical ‘messah’ or fat of the sacred dragon or crocodile. Note that we get the words Messiah and Moses from this; the Pharoah was hailed as being ‘born of’ as in the cases of Tuthmosis (born of Tuth or Thoth) and Ramesses (fashioned of Ra); so that there was an essential and divine dignity given. This points to a unique bloodline that we know as the blue-blood in the grail legends of Europe and from which also was the Hebrew House of David. David was descended also of this ‘mosis’, this divine essence of the sun. And of course, the exiled Pharoah, Akhenaten was hailed by his loyal adherents as the ‘mosis’. Even Oedipus is connected to these two other names of Akhenaten and Moses, and more directly does his story relate the mystery of the sexual and matrilineal mystery.
Further, studies of the Nencki Institute of Experimental Biology in Poland have produced Potassium Channel Openers that affect the Mitochondrial DNA in the hippocampus gland. And this Mitochondrial DNA has also been cloned as well. So there is a moral dilemma on the one hand. But on the other, there is an avenue of research that will not be broached nor even apparent to the mundane science of experimental biology. This is the purview of our Magick, that we can now begin to use this ancient knowledge and this modern science in order to move toward a modern approach to the procurement of the Philosopher’s Stone, the Summum Bonum, True Wisdom and Perfect Happiness.
There is a concept of Shakti-pat. that is, one person who has advanced, may in fact connect another, even though it will likely be temporary, to higher forms of realization, including Kundalini rising. Shakti pat means that water seeks it's own level. Anyone who is at a certain level, can energetically bring another person to that same level, if only temporarily, which it usually is only that…but it's enough to give the person a chance to see what they have to achieve to get there, and so is a form of initiation.
Because the Kundalini rising can make you think that you have "arrived" and that you know everything. it's a very dangerous time. It is not the end, but a beginning. The eastern texts talk about the Kundalini, the snake residing at the base of the spine, which when it rises, gives knowledge of all. Gilgamesh, after a soiree with a woman found this knowledge. Mohammed had the same problem, and countless others. It is suggested that one be very careful in the work. Kundalini is not to be played with at all, or mocked in arrogance. You can get really sick, even if you are dedicated. We simply cannot control everything. Kundalini is not a Goddess for nothing. She can resemble Kali in her devastation of the human psyche.
DNA and the Psyche
We communicate with our world through a previously unrecognized form of energy that operates outside the bounds of measured time and space; through a web or matrix of energy–the quantum- or non-local hologram. The Hopi tribe of Native American Peoples referred to it as the ‘Web of Spider Grandmother’ and even have a prophecy related to a giant web in the sky connecting the earth.
The Institute of Heart Math in Boulder Creek, CA, are pioneers in innovative heart-based technologies. In early '90s they found that every human heart is surrounded by a doughnut-shaped electromagnetic field between 5-8 ft. in diameter (probably much bigger, but machine could only measure that far). This is an acknowledged fact (JAMA & other profession journals).
When people are joined together in a "field" of shared emotion (eg. Pearl Harbor, Challenger disaster, JFK's death, 9-11 et al) there are measurable social effects (fewer traffic accidents, fewer emergency room admissions, fewer violent crimes, stock market rises, etc.) that disappear when the shared moment is over.
Our DNA directly influences our physical world through a form of energy that has been previously overlooked. There is a 5th nucleotide base in our DNA at birth that disappears after a short period of time, and so is rarely mentioned in the literature. As we begin to lose this 5th base, we begin aging & breakdown of bodily functions.
In 1991, a Russian physicist created a vacuum in a glass tube and shot photons into it, which scattered randomly, as expected. Then he put some human DNA into the vacuum and shot photons in. The photons lined up following the pattern of DNA. They took the DNA out of the vacuum and sent photons back in. These still lined up, even without the DNA's physical presence. This experiment was called "The DNA Phantom Effect" and has been replicated several times.
We influence our physical DNA through a non-physical form of energy that is directly linked to a certain modality of prayer and to human emotion. DNA is directly linked to and modulated by human emotion.
Quantum experiments are beginning to show that we have direct access to the four fundamental forces of nature in our bodies through our emotions. These four forces are electromagnetism, gravity, the strong nuclear force and the weak nuclear force. By changing our emotions we can directly affect these forces. This suggests the idea that when we fully activate a certain quality of emotion in our bodies, we are no longer mere observers of the events in the world and our bodies, but become participants and co-creators.
We are present in every one of the 10-11 dimensions of string theory, though not necessarily conscious in all the dimensions. "Ghosts" may be evidence of "non-local hologram/non-local energy" now being discussed by quantum physicists–existing in another dimension but lacking physical vehicle in our 3. Shroud of Turin left a mystery with quantum implications. Man in shroud accessed the 4 forces of creation to make the imprint.
We are directly related to the physical matter in the world around us, and DNA is the point of interaction between our selves & the world. In the Isaiah scrolls, Isaiah says (in effect) that there are many worlds and many possibilities unfolding at every moment, and through our feelings, we determine which of those play out in our world. When we feel the feelings in our body, that's what awakens the quantum possibility. Human emotion is the language that speaks to the forces of creation and awakens those possibilities, and that is what was edited out of the biblical texts. Certainly by the 4th c., we had lost that wisdom, and had become a very logic-based, non-"touchy-feely", male dominated, schematically oriented society. Quantum physics is now verifying the concepts the ancients tried to preserve.
In 1993, a U.S. Army Intelligence & Security Command research lab scraped tissue from inside of mouth of donor to obtain DNA. They then placed the donor in one room of building with his DNA in another room of building. The donor was then exposed to very powerful visual stimuli intended to exact a strong emotional response. With the donor’s emotional reaction, certain changes in the DNA were also observed to occur simultaneously. They separated donor and DNA by increasing distances between them, up to 50 miles of separation, and there was still a simultaneous reaction that clearly suggests a direct connection between the psyche and one’s DNA, even if removed from the body. Indeed, there are many well-documented instances of organ transplant recipients picking up on "inexplicable" and "miraculous" knowledge about the donors whose organs (and DNA) they received..
Of the 64 codes in DNA, about two-thirds of them seem to be dormant. This was thought to be a fixed state, but it now looks as though they can be activated willfully through an internal technology of thought, feeling and emotion. As well, they seem to become active in response to environmental conditions. In 1998 an AIDS research center did a global blood study and found that individuals who had tested positive for the AIDS virus had actually *turned on* 5 sets of genes that gave them up to 3000 times the normal immune response. Verified by U of Alabama.
Researchers have found that human emotion actually changes the physical shape of the double-helix of DNA. The very positive emotions relaxed the DNA to the point where the 2 strands were no longer intertwined. The very negative emotions wound the strands so tightly they were almost knotted. The "unused" DNA codes seem to be turned on or off based on the degree of winding of the strands. The positive emotions seem to activate the unused codes, negative emotions not only can't influence these "off" codes, but seem to have an adverse affect on even the permanently "on" codes.
There has in Magick, been developed, the theory of the ‘Aether’ that is correlated in science by the terms ‘non-local quantum hologram’ and "matrix of creation". So modern science is now recognizing that consciousness is a field of energy. As per the mystics of time immemorial, it seems that we are a very unique species with the power to directly interact with the forces of creation.
I. Embryology
THE TERM Embryology, in its widest sense, is applied to the various changes which take place during the growth of an animal from the egg to the adult condition: it is, however, usually restricted to the phenomena which occur before birth. Embryology may be studied from two aspects: (1) that of ontogeny, which deals only with the development of the individual; and (2) that of phylogeny, which concerns itself with the evolutionary history of the animal kingdom.
In vertebrate animals the development of a new being can only take place when a female germ cell or ovum has been fertilized by a male germ cell or spermatozoön. The ovum is a nucleated cell, and all the complicated changes by which the various tissues and organs of the body are formed from it, after it has been fertilized, are the result of two general processes, viz., segmentation and differentiation of cells. Thus, the fertilized ovum undergoes repeated segmentation into a number of cells which at first closely resemble one another, but are, sooner or later, differentiated into two groups: (1) somatic cells, the function of which is to build up the various tissues of the body; and (2) germinal cells, which become imbedded in the sexual glands—the ovaries in the female and the testes in the male—and are destined for the perpetuation of the species.
Having regard to the main purpose of this work, it is impossible, in the space available in this section, to describe fully, or illustrate adequately, all the phenomena which occur in the different stages of the development of the human body. Only the principal facts are given, and the student is referred for further details to one or other of the text-books 1 on human embryology.
1. The Animal Cell
All the tissues and organs of the body originate from a microscopic structure (the fertilized ovum), which consists of a soft jelly-like material enclosed in a membrane and containing a vesicle or small spherical body inside which are one or more denser spots. This may be regarded as a complete cell. All the solid tissues consist largely of cells essentially similar to it in nature but differing in external form.
In the higher organisms a cell may be defined as “a nucleated mass of protoplasm of microscopic size.” Its two essentials, therefore, are: a soft jelly-like material, similar to that found in the ovum, and usually styled cytoplasm, and a small spherical body imbedded in it, and termed a nucleus. Some of the unicellular protozoa contain no nuclei but granular particles which, like true nuclei, stain with basic dyes. The other constituents of the ovum, viz., its limiting membrane and the denser spot contained in the nucleus, called the nucleolus, are not essential to the type cell, and in fact many cells exist without them.
Cytoplasm (protoplasm) is a material probably of variable constitution during life, but yielding on its disintegration bodies chiefly of proteid nature. Lecithin and cholesterin are constantly found in it, as well as inorganic salts, chief among which are the phosphates and chlorides of potassium, sodium, and calcium. It is of a semifluid, viscid consistence, and probably colloidal in nature. The living cytoplasm appears to consist of a homogeneous and structureless ground-substance in which are embedded granules of various types. The mitochondria are the most constant type of granule and vary in form from granules to rods and threads. Their function is unknown. Some of the granules are proteid in nature and probably essential constituents; others are fat, glycogen, or pigment granules, and are regarded as adventitious material taken in from without, and hence are styled cell-inclusions or paraplasm. When, however, cells have been “fixed” by reagents a fibrillar or granular appearance can often be made out under a high power of the microscope. The fibrils are usually arranged in a network or reticulum, to which the term spongioplasm is applied, the clear substance in the meshes being termed hyaloplasm. The size and shape of the meshes of the spongioplasm vary in different cells and in different parts of the same cell. The relative amounts of spongioplasm and hyaloplasm also vary in different cells, the latter preponderating in the young cell and the former increasing at the expense of the hyaloplasm as the cell grows. Such appearances in fixed cells are no indication whatsoever of the existence of similar structures in the living, although there must have been something in the living cell to give rise to the fixed structures. The peripheral layer of a cell is in all cases modified, either by the formation of a definite cell membrane as in the ovum, or more frequently in the case of animal cells, by a transformation, probably chemical in nature, which is only recognizable by the fact that the surface of the cell behaves as a semipermeable membrane.
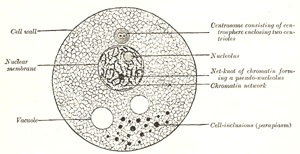
FIG. 1– Diagram of a cell. (Modified from Wilson.)
Nucleus.—The nucleus is a minute body, imbedded in the protoplasm, and usually of a spherical or oval form, its size having little relation to that of the cell. It is surrounded by a well-defined wall, the nuclear membrane; this encloses the nuclear substance (nuclear matrix), which is composed of a homogeneous material in which is usually embedded one or two nucleoli. In fixed cells the nucleus seems to consist of a clear substance or karyoplasm and a network or karyomitome. The former is probably of the same nature as the hyaloplasm of the cell, but the latter, which forms also the wall of the nucleus, differs from the spongioplasm of the cell substance. It consists of fibers or filaments arranged in a reticular manner. These filaments are composed of a homogeneous material known as linin, which stains with acid dyes and contains embedded in its substance particles which have a strong affinity for basic dyes. These basophil granules have been named chromatin or basichromatin and owe their staining properties to the presence of nucleic acid. Within the nuclear matrix are one or more highly refracting bodies, termed nucleoli, connected with the nuclear membrane by the nuclear filaments. They are regarded as being of two kinds. Some are mere local condensations (“net-knots”) of the chromatin; these are irregular in shape and are termed pseudo-nucleoli; others are distinct bodies differing from the pseudo-nucleoli both in nature and chemical composition; they may be termed true nucleoli, and are usually found in resting cells. The true nucleoli are oxyphil, i.e., they stain with acid dyes.
Most living cells contain, in addition to their protoplasm and nucleus, a small particle which usually lies near the nucleus and is termed the centrosome. In the middle of the centrosome is a minute body called the centriole, and surrounding this is a clear spherical mass known as the centrosphere. The protoplasm surrounding the centrosphere is frequently arranged in radiating fibrillar rows of granules, forming what is termed the attraction sphere.
Reproduction of Cells.—Reproduction of cells is effected either by direct or by indirect division. In reproduction by direct division the nucleus becomes constricted in its center, assuming an hour-glass shape, and then divides into two. This is followed by a cleavage or division of the whole protoplasmic mass of the cell; and thus two daughter cells are formed, each containing a nucleus. These daughter cells are at first smaller than the original mother cell; but they grow, and the process may be repeated in them, so that multiplication may take place rapidly. Indirect division or karyokinesis (karyomitosis) has been observed in all the tissues—generative cells, epithelial tissue, connective tissue, muscular tissue, and nerve tissue. It is possible that cell division may always take place by the indirect method.
The process of indirect cell division is characterized by a series of complex changes in the nucleus, leading to its subdivision; this is followed by cleavage of the cell protoplasm. Starting with the nucleus in the quiescent or resting stage, these changes may be briefly grouped under the four following phases (Fig. 2).
1. Prophase.—The nuclear network of chromatin filaments assumes the form of a twisted skein or spirem, while the nuclear membrane and nucleolus disappear. The convoluted skein of chromatin divides into a definite number of V-shaped segments or chromosomes. The number of chromosomes varies in different animals, but is constant for all the cells in an animal of any given species; in man the number is given by Flemming and Duesberg as twenty-four. 2 Coincidently with or preceding these changes the centriole, which usually lies by the side of the nucleus, undergoes subdivision, and the two resulting centrioles, each surrounded by a centrosphere, are seen to be connected by a spindle of delicate achromatic fibers the achromatic spindle. The centrioles move away from each other—one toward either extremity of the nucleus—and the fibrils of the achromatic spindle are correspondingly lengthened. A line encircling the spindle midway between its extremities or poles is named the equator, and around this the V-shaped chromosomes arrange themselves in the form of a star, thus constituting the mother star or monaster.
2. Metaphase.—Each V-shaped chromosome now undergoes longitudinal cleavage into two equal parts or daughter chromosomes, the cleavage commencing at the apex of the V and extending along its divergent limbs.
3. Anaphase.—The daughter chromosomes, thus separated, travel in opposite directions along the fibrils of the achromatic spindle toward the centrioles, around which they group themselves, and thus two star-like figures are formed, one at either pole of the achromatic spindle. This constitutes the diaster. The daughter chromosomes now arrange themselves into a skein or spirem, and eventually form the network of chromatin which is characteristic of the resting nucleus.
4. Telophase.—The cell protoplasm begins to appear constricted around the equator of the achromatic spindle, where double rows of granules are also sometimes seen. The constriction deepens and the original cell gradually becomes divided into two new cells, each with its own nucleus and centrosome, which assume the ordinary positions occupied by such structures in the resting stage. The nuclear membrane and nucleolus are also differentiated during this phase.
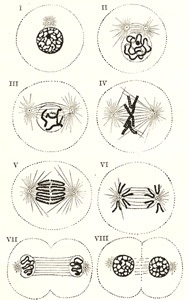
FIG. 2– Diagram showing the changes which occur in the centrosomes and nucleus of a cell in the process of mitotic division. (Schäfer.) I to III, prophase; IV, metaphase; V and VI, anaphase; VII and VIII, telophase.
Note 1. Manual of Human Embryology, Keibel and Mall; Handbuch der vergleichenden und experimentellen Entwickelungslehre der Wirbeltiere, Oskar Hertwig; Lehrbuch der Entwickelungsgeschichte, Bonnet; The Physiology of Reproduction, Marshall. [back]
Note 2. Dr. J. Duesberg, Anat. Anz., Band xxviii, S. 475.
2. The Ovum
The ova are developed from the primitive germ cells which are imbedded in the substance of the ovaries. Each primitive germ cell gives rise, by repeated divisions, to a number of smaller cells termed oögonia, from which the ova or primary oöcytes are developed.
Human ova are extremely minute, measuring about 0.2 mm. in diameter, and are enclosed within the egg follicles of the ovaries; as a rule each follicle contains a single ovum, but sometimes two or more are present. 3 By the enlargement and subsequent rupture of a follicle at the surface of the ovary, an ovum is liberated and conveyed by the uterine tube to the cavity of the uterus. Unless it be fertilized it undergoes no further development and is discharged from the uterus, but if fertilization take place it is retained within the uterus and is developed into a new being.
In appearance and structure the ovum (Fig. 3) differs little from an ordinary cell, but distinctive names have been applied to its several parts; thus, the cell substance is known as the yolk or oöplasm, the nucleus as the germinal vesicle, and the nucleolus as the germinal spot. The ovum is enclosed within a thick, transparent envelope, the zona striata or zona pellucida, adhering to the outer surface of which are several layers of cells, derived from those of the follicle and collectively constituting the corona radiata.

FIG. 3– Human ovum examined fresh in the liquor folliculi. (Waldeyer.) The zona pellucida is seen as a thick clear girdle surrounded by the cells of the corona radiata. The egg itself shows a central granular deutoplasmic area and a peripheral clear layer, and encloses the germinal vesicle, in which is seen the germinal spot.
Yolk.—The yolk comprises (1) the cytoplasm of the ordinary animal cell with its spongioplasm and hyaloplasm; this is frequently termed the formative yolk; (2) the nutritive yolk or deutoplasm, which consists of numerous rounded granules of fatty and albuminoid substances imbedded in the cytoplasm. In the mammalian ovum the nutritive yolk is extremely small in amount, and is of service in nourishing the embryo in the early stages of its development only, whereas in the egg of the bird there is sufficient to supply the chick with nutriment throughout the whole period of incubation. The nutritive yolk not only varies in amount, but in its mode of distribution within the egg; thus, in some animals it is almost uniformly distributed throughout the cytoplasm; in some it is centrally placed and is surrounded by the cytoplasm; in others it is accumulated at the lower pole of the ovum, while the cytoplasm occupies the upper pole. A centrosome and centriole are present and lie in the immediate neighborhood of the nucleus.
Germinal Vesicle.—The germinal vesicle or nucleus is a large spherical body which at first occupies a nearly central position, but becomes eccentric as the growth of the ovum proceeds. Its structure is that of an ordinary cell-nucleus, viz., it consists of a reticulum or karyomitome, the meshes of which are filled with karyoplasm, while connected with, or imbedded in, the reticulum are a number of chromatin masses or chromosomes, which may present the appearance of a skein or may assume the form of rods or loops. The nucleus is enclosed by a delicate nuclear membrane, and contains in its interior a well-defined nucleolus or germinal spot.
Coverings of the Ovum.—The zona striata or zona pellucida (Fig. 3) is a thick membrane, which, under the higher powers of the microscope, is seen to be radially striated. It persists for some time after fertilization has occurred, and may serve for protection during the earlier stages of segmentation. It is not yet determined whether the zona striata is a product of the cytoplasm of the ovum or of the cells of the corona radiata, or both.
The corona radiata (Fig. 3) consists or two or three strata of cells; they are derived from the cells of the follicle, and adhere to the outer surface of the zona striata when the ovum is set free from the follicle; the cells are radially arranged around the zona, those of the innermost layer being columnar in shape. The cells of the corona radiata soon disappear; in some animals they secrete, or are replaced by, a layer of adhesive protein, which may assist in protecting and nourishing the ovum.
The phenomena attending the discharge of the ova from the follicles belong more to the ordinary functions of the ovary than to the general subject of embryology, and are therefore described with the anatomy of the ovaries.
Maturation of the Ovum.—Before an ovum can be fertilized it must undergo a process of maturation or ripening. This takes place previous to or immediately after its escape from the follicle, and consists essentially of an unequal subdivision of the ovum (Fig. 4) first into two and then into four cells. Three of the four cells are small, incapable of further development, and are termed polar bodies or polocytes, while the fourth is large, and constitutes the mature ovum. The process of maturation has not been observed in the human ovum, but has been carefully studied in the ova of some of the lower animals, to which the following description applies.
It was pointed out on page 37 that the number of chromosomes found in the nucleus is constant for all the cells in an animal of any given species, and that in man the number is probably twenty-four. This applies not only to the somatic cells but to the primitive ova and their descendants. For the purpose of illustrating the process of maturation a species may be taken in which the number of nuclear chromosomes is four (Fig. 5). If an ovum from such be observed at the beginning of the maturation process it will be seen that the number of its chromosomes is apparently reduced to two. In reality, however, the number is doubled, since each chromosome consists of four granules grouped to form a tetrad. During the metaphase (see page 37) each tetrad divides into two dyads, which are equally distributed between the nuclei of the two cells formed by the first division of the ovum. One of the cells is almost as large as the original ovum, and is named the secondary oöcyte; the other is small, and is termed the first polar body. The secondary oöcyte now undergoes subdivision, during which each dyad divides and contributes a single chromosome to the nucleus of each of the two resulting cells.
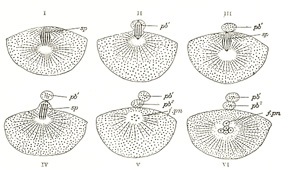
FIG. 4– Formation of polar bodies in Asterias glacialis. (Slightly modified from Hertwig.) In I the polar spindle (sp) has advanced to the surface of the egg. In II a small elevation (pb1) is formed which receives half of the spindle. In III the elevation is constricted off, forming the first polar body (pb1), and a second spindle is formed. In IV is seen a second elevation which in V has been constricted off as the second polar body (pb2). Out of the remainder of the spindle (f.pn in VI) the female pronucleus is developed.
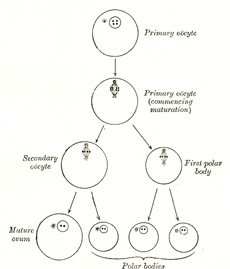
FIG. 5– Diagram showing the reduction in number of the chromosomes in the process of maturation of the ovum.
This second division is also unequal, producing a large cell which constitutes the mature ovum, and a small cell, the second polar body. The first polar body frequently divides while the second is being formed, and as a final result four cells are produced, viz., the mature ovum and three polar bodies, each of which contains two chromosomes, i.e., one-half the number present in the nuclei of the somatic cells of members of the same species. The nucleus of the mature ovum is termed the female pronucleus.
Note 3. See description of the ovary on a future page.
Note 4. See description of the ovary on a future page.
3. The Spermatozoön
The spermatozoa or male germ cells are developed in the testes and are present in enormous numbers in the seminal fluid. Each consists of a small but greatly modified cell. The human spermatozoön possesses a head, a neck, a connecting piece or body, and a tail (Fig. 6).
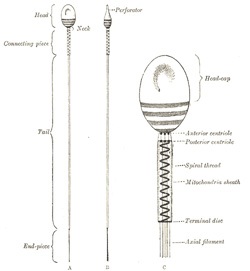
FIG. 6– Human spermatozoön. Diagrammatic. A. Surface view. B. Profile view. In C the head, neck, and connecting piece are more highly magnified. (See enlarged image)
The head is oval or elliptical, but flattened, so that when viewed in profile it is pear-shaped. Its anterior two-thirds are covered by a layer of modified protoplasm, which is named the head-cap. This, in some animals, e. g., the salamander, is prolonged into a barbed spear-like process or perforator, which probably facilitates the entrance of the spermatozoön into the ovum. The posterior part of the head exhibits an affinity for certain reagents, and presents a transversely striated appearance, being crossed by three or four dark bands. In some animals a central rodlike filament extends forward for about two-thirds of the length of the head, while in others a rounded body is seen near its center. The head contains a mass of chromatin, and is generally regarded as the nucleus of the cell surrounded by a thin envelope.
The neck is less constricted in the human spermatozoön than in those of some of the lower animals. The anterior centriole, represented by two or three rounded particles, is situated at the junction of the head and neck, and behind it is a band of homogeneous substance.
The connecting piece or body is rod-like, and is limited behind by a terminal disk. The posterior centriole is placed at the junction of the body and neck and, like the anterior, consists of two or three rounded particles. From this centriole an axial filament, surrounded by a sheath, runs backward through the body and tail. In the body the sheath of the axial filament is encircled by a spiral thread, around which is an envelope containing mitochondria granules, and termed the mitochondria sheath.
The tail is of great length, and consists of the axial thread or filament, surrounded by its sheath, which may contain a spiral thread or may present a striated appearance. The terminal portion or end-piece of the tail consists of the axial filament only.
FIG. 7– Scheme showing analogies in the process of maturation of the ovum and the development of the spermatids (young spermatozoa). (See enlarged image)
Krause gives the length of the human spermatozoön as between 52μ and 62μ, the head measuring 4 to 5μ, the connecting piece 6μ, and the tail from 41μ to 52μ.
By virtue of their tails, which act as propellers, the spermatozoa are capable of free movement, and if placed in favorable surroundings, e. g., in the female passages, will retain their vitality and power of fertilizing for several days. In certain animals, e. g., bats, it has been proved that spermatozoa retained in the female passages for several months are capable of fertilizing. 7
The spermatozoa are developed from the primitive germ cells which have become imbedded in the testes, and the stages of their development are very similar to those of the maturation of the ovum. The primary germ cells undergo division and produce a number of cells termed spermatogonia, and from these the primary spermatocytes are derived. Each primary spermatocyte divides into two secondary spermatocytes, and each secondary spermatocyte into two spermatids or young spermatozoa; from this it will be seen that a primary spermatocyte gives rise to four spermatozoa. On comparing this process with that of the maturation of the ovum (Fig. 7) it will be observed that the primary spermatocyte gives rise to two cells, the secondary spermatocytes, and the primary oöcyte to two cells, the secondary oöcyte and the first polar body. Again, the two secondary spermatocytes by their subdivision give origin to four spermatozoa, and the secondary oöcyte and first polar body to four cells, the mature ovum and three polar bodies. In the development of the spermatozoa, as in the maturation of the ovum, there is a reduction of the nuclear chromosomes to one-half of those present in the primary spermatocyte. But here the similarity ends, for it must be noted that the four spermatozoa are of equal size, and each is capable of fertilizing a mature ovum, whereas the three polar bodies are not only very much smaller than the mature ovum but are incapable of further development, and may be regarded as abortive ova.
4. Fertilization of the Ovum
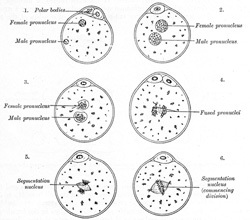
FIG. 8– The process of fertilization in the ovum of a mouse. (After Sobotta.) (See enlarged image)
Fertilization consists in the union of the spermatozoön with the mature ovum (Fig. 8). Nothing is known regarding the fertilization of the human ovum, but the various stages of the process have been studied in other mammals, and from the knowledge so obtained it is believed that fertilization of the human ovum takes place in the lateral or ampullary part of the uterine tube, and the ovum is then conveyed along the tube to the cavity of the uterus—a journey probably occupying seven or eight days and during which the ovum loses its corona radiata and zona striata and undergoes segmentation. Sometimes the fertilized ovum is arrested in the uterine tube, and there undergoes development, giving rise to a tubal pregnancy; or it may fall into the abdominal cavity and produce an abdominal pregnancy. Occasionally the ovum is not expelled from the follicle when the latter ruptures, but is fertilized within the follicle and produces what is known as an ovarian pregnancy. Under normal conditions only one spermatozoön enters the yolk and takes part in the process of fertilization. At the point where the spermatozoön is about to pierce, the yolk is drawn out into a conical elevation, termed the cone of attraction. As soon as the spermatozoön has entered the yolk, the peripheral portion of the latter is transformed into a membrane, the vitelline membrane which prevents the passage of additional spermatozoa. Occasionally a second spermatozoön may enter the yolk, thus giving rise to a condition of polyspermy: when this occurs the ovum usually develops in an abnormal manner and gives rise to a monstrosity. Having pierced the yolk, the spermatozoön loses its tail, while its head and connecting piece assume the form of a nucleus containing a cluster of chromosomes. This constitutes the male pronucleus, and associated with it there are a centriole and centrosome. The male pronucleus passes more deeply into the yolk, and coincidently with this the granules of the cytoplasm surrounding it become radially arranged. The male and female pronuclei migrate toward each other, and, meeting near the center of the yolk, fuse to form a new nucleus, the segmentation nucleus, which therefore contains both male and female nuclear substance; the former transmits the individualities of the male ancestors, the latter those of the female ancestors, to the future embryo. By the union of the male and female pronuclei the number of chromosomes is restored to that which is present in the nuclei of the somatic cells.
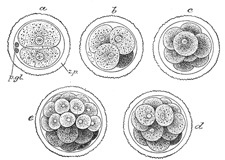
FIG. 9– First stages of segmentation of a mammalian ovum. Semidiagrammatic. (From a drawing by Allen Thomson.) z.p. Zona striata. p.gl. Polar bodies. a. Two-cell stage. b. Four-cell stage. c. Eight-cell stage. d, e. Morula stage.
THE FEMALE: FIRST FORM
& SOURCE OF LIFE
RESCRÎPTUM ATHENADORÎ Nº 2: IN CULTÛS PRÆNÔTIÔNEM MULIERIBUS
RESCRIPT Nº 2 OF ATHENADORUS: AGAINST RELIGIOUS PREJUDICE TOWARD WOMEN
The male produces the seed complete: the female merely broods his creation till it be of a size survivable. For having enticed him into sin, it is her eternal punishment to have to carry the children he produces and to deliver them to him in pain and anguish.
The female is a weak and empty vessel until the male bestow his fertility upon her. The male was the original form created, the female was created secondarily from him, as a divine afterthought, to be his helper, his servant and upon whom he might beget his sons.
Thus is the nature of reproduction portrayed in the mythology of the father-god cults. It is incorrect. It is wrong. It is damnably false.
All living things begin their development as immature females of their species. No matter what DNA mix it carries, every fetus begins its life with the form of the female. The female is the original, not the male.
It is easy to see why the error of male pre-eminence arose. It is the male that produces a substance external and unusual, clearly associated with reproduction, whereas the female generates no such obvious material: the female appears an empty vessel. From the male’s production of that visible substance, the analogy arose that the male was doing exactly that act performed by farmers, that is, he was placing fully viable seed into a temporary, enclosing environment for its intermediate development. But this analogy is false, absolutely false.
That substance visibly contributed by the male transfers half of the DNA necessary for the full functioning of the nuclear DNA of the cells that will become a new individual of its species. This one-half of the nuclear DNA is the entire contribution the male makes to the physical conception of this new life. It is vitally important, yet the emphasis placed on it as the end-all, be-all of procreation has been so profoundly overdone as to have become absurd–it has perverted the laws, attitudes and structures of human society.
The contribution the female makes to this new life is far greater than that allotted to the male. In addition to the remaining half of the nuclear DNA, there are several structures, processes and chemicals that the female builds into the egg, thus into the cells, of the new individual. These objects are wholly contributed by the female. They never come from the male parent: it is physically impossible.
The egg is, in fact, a cell of the mother’s body, a cell from which half of the nuclear DNA has been omitted during its process of becoming an egg cell. In its internal organization, messages, structures, and functions, it is exactly like the other cells of the mother. Among these maternal substances, objects and processes, the most important are the mitochondria; the ribosomes and their associated RNA’s; and disease-fighting chemicals, the immuno-globulins.1
The fetus is bathed in the immuno-globulins suffused into the womb from the fluids of the mother’s body. These substances persist for some time in the newborn, as its initial, primary disease-fighting capacity.
The maternal ribosomes and RNA’s made available to the embryo allow it to grow until it has reached a size and complexity that permit it to replace them with those of its own production.
However, of those specific three inherited from the mother, the most persistently important are the mitochondria. The mitochondria convert the energy contained in the chemical bonds of glucose molecules into an intermediate form that can be safely and easily moved about the cell itself, then released when and where needed to power those chemical reactions necessary for the life of the cell.
Mitochondria contain their own DNA, DNA unrelated to that located in the nucleus of the cell. This non-nuclear, genetic material controls the nature and efficiency of the chemical reactions mediated by the mitochondria, as well as the reproduction of the mitochondria themselves. The DNA of the cell’s nucleus contains no instructions for making mitochondria: only a mitochondrion can produce mitochondria.
Those mitochondria bestowed by the mother are the only ones the child will ever receive. They will replicate themselves with every cell division until they are contained in each living cell of the child’s body, to power all tasks necessary in the body. And they will continue this replication and empowerment throughout the child’s existence.
Life as we know it is impossible without mitochondria. Yet all mitochondria are inherited from the mother and only from the mother.
The sexual form of an individual is determined by its sex chromosomes. For mammals in particular, two X-chromosomes are present in the female; one X-chromosome and a Y-chromosome exist in the male.
However, in females one of those two X-chromosomes is non-functional; it shrivels into what is called a Barr body. Because of this, females are actually creatures of one X-chromosome. Males are also creatures with only one X-chromosome. Their Y-chromosome (that takes the “place” of the Barr body) causes few processes other than those necessary to change the original one-X-chromosome form—the female—into that other, one-X-chromosome form, the form that we recognize as the male.
The Y-chromosome is that which was added by evolutionary processes to create the male from the female.
While the XY-chromosome fetus is in the womb it will, under normal circumstances, undergo two surges of hormonal activity that result in the development of the fully formed XY-individual. These chemical fluxes do not occur in the development of the female, the XX-individual: they exist only in the maturation process of the male fetus.
The first set of hormones stops the production of fully-formed female sexual characteristics. This is required because the functional X-chromosome of both males and females contains complete instructions for molding the fetus into a female, a process that will continue automatically if not interrupted. To produce a ‘canonical’ male form, these genes must be switched off, by the chemical action of hormones, before the second surge of chemicals is allowed to arise.
That second set of hormones sets in motion the production of male sexual characteristics. The existence of these two hormonal surges explains the existence among normal-chromosome’d individuals of sexual neuters, hermaphrodites, Y-chromosome-females and regularly formed individuals. In hermaphrodites, the first surge does not occur; in neuters, the second surge does not occur; in Y-chromosome-females, neither surge occurs.
The male never loses those female structures and characteristics it starts out with, the most obvious example being his nipples. All male sexual characteristics are derived from female structures: the male has no parts that do not exist in their original form in the female.
Many young men, while watching the changes worked on them by the metamorphosis of puberty, discover a ‘suture’ line running along the center axis of the scrotum, continuing up along the underside of the penis.2 They do not realize that this ‘scar’ is the line along which what had been their original, external female organs came together to form these structures. That suture line marks where their labia fused in the process of forming the penis and the scrotum. Contrary to the biblically derived idea that the female is a male with no penis, the truth is actually that the male is a female with an adapted vulva.
When that second surge of hormones flows through the body of the XY-fetus, those structures that had just previously been made to stop developing into ovaries start to descend from their original ovary-positions, down and out through the body wall into the pouch the body has simultaneously been creating from part of what had been the fetus’s labia.
The descent of the gonads from their ovary-positions into the developing scrotum is inherently a destructive process. It requires that the incipient testicles push their way through the musculo-tendinous membranes of the body cavity to reach the outside. Because these membranes have already formed in a way that is female-specific in how they reinforce and support the internal organs, this pushing of the testicles through them creates weaknesses in those membranes.3 This is the reason males can so easily develop hernias in this area: the external testicles are, in fact, already a herniation.
In retelling the two myths that he combined to create the biblical myth of Noah, the editor of that text has Yavah commanding Noah to take, in one verse,5 two of each animal–one male and one female, and in other verses,6 seven pairs of each “clean” animal and two of each “unclean”, of each beast “the male and his female”. Among other observations,7 it is very interesting that this creator god has forgotten that there are several species of animals–species that he is supposed to have created specifically and separately–in which there are no males.8 In these species, the females produce eggs that are already fertile–no male input is necessary, nor, indeed, is it possible, for there are no males in these species.
There are several species of fish in which all offspring are born female. On the occurrence of certain environmental stimuli, some of the females of these species will, from hormonal actions, turn into males.
Although there are fish species that bear hermaphroditic offspring, capable of shifting between male-functionality and female-functionality, there are no species, fish or otherwise, that produce only male offspring.
The female is the original form of life: the male is the second form, generated from that original form in each instance.
The mythology that holds that the male is the original, true form has been used for many thousands of years as false evidence of the divinely ordained superiority of the male. It has been cited as legitimization of man’s abuse of woman. From denial of rights to rape to murder, this lie has been perpetrated against her.
Because man thought woman to be incompletely developed, imperfect in body, mind and spirit, she has long been treated as inferior. Few realize today that up until around a hundred years ago, in many parts of the United States a woman could not own property in her own name. In most regions, all property a young woman’s father held for her was transferred, at marriage, not to her, but to her husband. If her husband died, her eldest son took her in tutelage. Thus she was passed from male domination to male domination.
The assumption of male preeminence has caused the work of woman to be undervalued, her role in society to be ignored, her effects on history to be dismissed.
Because the male is merely a form derived from the female original, he is indeed still female. Male violence against women is a depraved expression of his own, societally inflicted self-hatred.
This perverted condition arose in its most heinous form and enforcement in the late days of the Roman Empire. Having begun with Constantine’s imposition of a misogynist, gynophobic religion, constantly reinforced throughout the Middle Ages, and continuing to this day, the disempowerment of woman has been horribly, viciously, violently enforced. This violence must now end.
The male is not superior to the female, nor is she superior to him. She is the source. He is its companion. She is the font of the great Waters of Life. He is the one whose swimming helps keep those waters alive. We are all one species.
Sexuality is a continuum, an arch o’er-reaching the sexes. It unites all peoples in our one, full purpose: the peaceful enrichment of the burgeoning lives of all humans.
We are each gifted as individuals, endowed by our birth with unique skills and strengths. Yet these gifts of the Earth, our ultimate mother, cannot be fully and freely made use of until we are all free.
That heinous misunderstanding, the sanctified vilification of the female is unnatural. That too-long list of acts of bias, of bigotry, of violence against women cries out, the unsilenceable witness of terror perpetuated by fear, by greed and by ignorance. We must commit ourselves to removing that ignorance, to quenching that greed, to ending that fear.
All attitudes, all doctrines, all institutions that propagate this mistake of male primacy, who perpetuate the perversion of male-specific creation, birth or re-birth, must yield to the truth of the female source, or be destroyed by the truth.
As the female is the true, original form of life, Woman’s Rights are the bulwark, the source, the very foundation of all Human Rights. Only when Woman is allowed Her full measure of Freedom will Her Children, all Her People, be vouchsafed their true liberty.
ATHENADORUS per Danaan Press, EY 5.3018, Copyright © 1996
www.danaanpress.com
¬ 1Athenadorus, The Athenadoran Library; The Danaa §VI, Items 1-3. Danaan Press 1982, 1996.
¬ 2 This “suture-line” is separately formed from, though usually continuous with, the other obvious suture-like scar that goes from the genitalia to the anus, and which is present in some way on all vertebrates (and which is especially visible on hairless mammals). This line is formed earlier in the development of the embryo to seal the “bottom” end of the tube that becomes the digestive system.
¬ 3 This occurs quite late in fetal development: for humans, it is often not completed until the seventh month of gestation.
¬4The processes involved are essentially these (keeping in mind that all structure names should be prefixed with “proto-”, or “the developing …”):
• the ovaries and the fingers of the fallopian tubes (which lengthen with this motion) move forward and down, push through the body wall and descend in front of the symphysis pubis into the sack prepared by the ‘suturing’ of the external labia, i.e., the scrotum, where they are modified to become the testes and epididymes
• the fallopian tube and horn of each side of the uterus form the ductus deferens, or vas deferens, leading from that side’s testis and epididymis back into the body cavity, up, over and behind the bladder; each horn of the uterus also contributes to forming the seminal vesicle attached to the vas deferens on that side
• the vasa deferentia meet in the prostate, made from what had been forming as the body of the uterus, the cervix and vagina; within the prostate, the pathways leading to the outside are made to change from the female’s three-exit system to the male’s two-exit form by the joining of the ducti deferentia with the female’s ureter, creating the male’s combined pathway, the urethra
• the clitoris extends forward to form one of the internal structures of the penis, the corpus cavernosum, around which the internal labia have come together to form both the body of the penis, and the urethra passing through it, which extends to end in the glans (or ‘head’) of the penis; all of which is covered by layers of skin formed from the external labia
This description is a general, highly non-specific list of independent structural changes. These separate processes do not happen sequentially, but progress relatively in tandem.
¬ 5from the earlier, 40-day flood myth
¬ 6from the later, 1-year flood myth
¬ 7 such as the problem of how Noah would have gone about choosing which female was “his female” of the male of animals that are “polygamous”, such as lions, seals, walruses, or of animals that do not form permanent pairs (which is almost all).
¬ 8Species such as the whiptail lizards of the American Southwest.
¬ 9 For a more in-depth treatment of the complexities of the sex of fish, see “Fishy Sex; Uncovering the wild ways of fish” by Tina Adler, Science News, Vol. 148, No. 17, October 21, 1995, pp. 266-7.
[Available online at http://www.sciencenews.org/sn_edpik/ls_4.htm]